Selected Self-Organization and the Semiotics of Evolutionary Systems
Citation: Rocha, Luis M. [1998]."Selected Self-Organization
and the Semiotics of Evolutionary Systems". In: Evolutionary Systems: The Biological and Epistemological Perspectives on Selection
and Self- Organization, . S. Salthe, G. Van de Vijver, and M. Delpos (eds.). Kluwer
Academic Publishers, pp. 341-358.
This paper is available in Adobe Acrobat (.pdf) format. This html version may be missing some special symbols, which you can get with the pdf version.
Abstract: In this paper I sketch a rough taxonomy of self-organization which may be of
relevance in the
study of cognitive and biological systems. I frame the problem both in terms of the language of
second-
order cybernetics as well as the language of current theories of self-organization and complexity.
The
goal of establishing such a taxonomy is to allow for a classification of different tools used both in
Artificial Intelligence and Artificial Life, so that different aspects of cognitive and biological
systems may
be incorporated in more accurate models of such systems. In particular, I defend, on the one
hand, that
self-organization alone is not rich enough for our intended simulations, and on the other, that
genetic
selection in biology and symbolic representation in cognitive science alone leave out the very
important
(self-organizing) characteristics of particular embodiments of evolving and learning systems.
Keywords: Self-organization, Semantic Closure, Semiotics, Emergence, Evolutionary
Strategies,
Artificial Life, Artificial Intelligence.
1 Eigenbehavior and Emergent Representation
Heinz von Foerster [1965, 1969, 1977] equated the ability of an organization to classify its
environment
with the notion of eigenbehavior. He postulated the existence of some stable structures
(eigenvalues)
which are maintained in the operations of an organization's dynamics. Following Piaget [von
Foerster,
1977], he observed that any specific instance of observation of such an organization, will still be
the result
of an indefinite succession of cognitive/sensory-motor operations. This reiterated the
constructivist
position that observables do not refer directly to real world objects, but are instead the result of
an infinite
cascade of cognitive and sensory-motor operations in some environment/subject coupling.
Eigenvalues
are self-defining, or self-referent, through the imbedding dynamics _ implying a complementary
relationship (circularity, closure) between eigenvalues and cognitive/sensory-motor operators:
one
implies, or defines, the other. "Eigenvalues represent the externally observable
manifestations of the
(introspectively accessible) cognitive [operations]". [von Foerster, 1977, page 278, italics
added]. Further,
"Ontologically, Eigenvalues and objects, and likewise, ontogenetically, stable behavior and the
manifestation of a subject's 'grasp' of an object cannot be distinguished." [von Foerster, 1977,
page 280].
Eigenbehavior is thus used to define the behavior of autonomous, cognitive systems,
which through the
closure (self-referential recursion) of the sensory-motor interactions in their nervous systems,
give rise to
perceptual regularities as objects [Varela, 1979, chapter 13].
"Eigenvalues are discrete (even if the domain of [their observables] is continuous)".
In other words, even
if the domain of an observable is continuous, its cognitive representation through
cognitive/sensory-motor
operators into eigenvalues must be discrete. This is a result of the stability of eigenvalues in the
recursive
chain of cognitive operators, if an eigenvalue changes its structure, thus ending the frame of
stability, it
will either revert to unstable structures (varying at each cognitive operation), in which case the
eigenvalue
representation is lost, or form another frame of stability with a new eigenvalue representation.
Insummary, eigenvalues are discrete representations of observables maintained by the successive
cognitive
operations of a cognitive agent. Notice that the representations and their stability are specific to
the
particular cognitive operations and how they recognize observables, that is, these discrete
representations
exist only in relation to the very same operators that define them. Any system, cognitive or
biological,
which is able to relate internally, self-organized, stable structures (eigenvalues) to constant
aspects of its
own interaction with an environment can be said to observe eigenbehavior. Such systems are
defined as
organizationally closed because their stable internal states can only defined in terms of
the overall
dynamic structure that supports them. Organizationally closed systems are also
informationally open
[Pask, 1992], since they have the ability to classify their constructed environment in what might
be
referred to as emergent representation.
1.1 Attractor behavior, self-organization, and constructivism
An eigenvalue of an organizationally closed system can be seen as an attractor of a
self-organizing
dynamical system. The global "cooperation" of the elements of a dynamical system which
spontaneously
emerges when an attractor state is reached is understood as self-organization [von Foerster, 1960;
Haken,
1977; Prigogine, 1985; Forrest, 1991; Kauffman, 1993]. The attractor behavior of any dynamical
system
is dependent on the structural operations of the latter, e.g. the set of boolean functions in a
boolean
network. Speaking of an attractor makes sense only in relation to its dynamical system, likewise,
the
attractor landscape defines its corresponding dynamical system. Further, attractor values can be
used to
refer to observables accessible to the dynamical system in its environment and therefore perform
relevant
classifications in such environment (e.g. neural networks). Naturally, and this is the crux of the
constructivist position in the theory of organizationally closed systems, not all possible
distinctions in
some environment can be "grasped" by the autonomous system: it can only classify those aspects
of its
environment/sensory-motor/cognitive interaction which result in the maintenance of some
internally
stable state or attractor (eigenvalue). In other words, not everything "out there" is accessible; only
those
things that a particular physiology can construct with the stabilities of its own dynamics are. As
with
eigenvalues, attractors must be discrete even if used to refer to continuous observables.
1.2 Emergence and levels of description
There are three levels that need to be addressed when dealing with the notion of emergent
representation.
First, there is the material, dynamical, substrate, which will be the causal basis for all other levels
that we
may further distinguish. Secondly, we have the attractor behavior of this dynamics. Finally, we
have the
utilization of the set of attractors (eigenvalues) as referents for some aspects of the interaction of
the
dynamical system itself with its environment, that is, as tokens for eigenbehavior. This indirect,
constructed, "referring" results from the structural coupling [Maturana and Varela, 1987] of the
dynamical system with the environment, and can be understood as a semantic relation.
The level of eigenvalues is emergent to the dynamics because it cannot be explained solely by a
description of the latter. Stability of dynamical states is not expressed in the language of the
interactions
between the components of a dynamical system. At this lower level, there is no distinction
between a
stable and an unstable state. For instance, the transition rules of Conway's game of Life
cannot describe
what "blinkers" and "gliders" are. Likewise, the level of eigenbehaviors, or the function
of attractors as
referring to some constructed reality, is emergent to the eigenvalues since the latter can only
describe
stabilities of the dynamics and not any "standing for" relation necessary for eigenbehavior (e.g.
streams of
gliders as information carriers in a universal computer built out of Life patterns
[Poundstone, 1987]). No
physical or formal description of the dynamical system and its attractors alone will completely
explainthis functional dimension [see Rocha, 1994a, 1995b; Rosen, 1995; Pattee, 1995]. Hence,
we need
complementary descriptions of the several levels involved in such organizationally closed,
emergent,
systems [Pattee, 1978].
2 Embodiment and self-organization
Varela, Thompson, and Rosch [1991] have proposed an embodied, inclusive, approach to
cognition which
acknowledges the different levels of description necessary to effectively deal with emergent
representation, or in von Foerster's terms, eigenbehavior. Cognitive science used to be
traditionally
concerned solely with those aspects of cognitive representation which can be described as
symbolic. In
other words, it was concerned with the semantic relation between cognitive categories and their
environmental counterparts through some direct representational relation (intentionalty), without
taking
into account any sort of material or internal organizational constraints: real-world categories
directly
represented by discrete symbols which could be freely manipulated. The connectionist, emergent,
or self-
organizing paradigm has changed this focus to the lower level of attractor behavior. That is,
cognitive
systems are defined as those systems capable of self-organizing their components into discrete
basins of
attraction used to discriminate the environment they are able to construct. Classifications become
subsymbolic and reside in some stable pattern of activation of the dynamic system's components,
instead
of based on some higher level symbols (emergent representation).
2.1 Selected Self-organization: structural change and increasing variety
What is usually referred to as self-organization is the spontaneous formation of well
organized structures,
patterns, or behaviors, from random initial conditions. The systems used to study this
phenomenon are
referred to as dynamical systems: state-determined systems. They possess a large number of
elements or
variables, and thus very large state spaces. However, when started with some initial conditions
they tend
to converge to small areas of this space (attractor basins) which can be interpreted as a form of
self-
organization. Since such formal dynamical systems are usually used to model real dynamical
systems
such as chemical networks of reactions, non-equilibrium thermodynamic behavior [Nicolis and
Prigogine,
1977] the conclusion is that in nature, there is a tendency for spontaneous self-organization
which is
therefore universal [Kauffman, 1993].
This process of self-organization is also often interpreted as the evolution of order from a
disordered start.
Self-organizing approaches to life (biological or cognitive), in particular second-order
cybernetics [see
Pask, 1992], take chaotic attractors as the mechanism which will be able to increase the variety
(physiological or conceptual) of organizationally closed systems. External random perturbations
will lead
to internal chaotic state changes; the richness of strange attractors is converted to a wide variety
of
discriminative power. Dynamic systems such as boolean networks clearly have the ability to
discriminate
inputs. Generally, the attractors of their dynamics are used to represent events in their
environments:
depending on inputs, the network will converge to different attractors. However, for any
classification to
have survival value, it must relate its own constructed states (attractors) to relevant events in its
environment, thus, similar events in the world should correspond to the same attractor basin.
Chaotic
systems clearly do not have this property due to their sensitivity to initial conditions. Ordered
systems
follow this basic heuristic.
Kauffman [1993, page 232] further hypothesizes that "living systems exist in the [ordered]
regime near
the edge of chaos, and natural selection achieves and sustains such a poised state". This
hypothesis is
based on Packard's [1988] work showing that when natural selection algorithms are applied to
dynamic
systems, with the goal of achieving higher discriminative power, the parameters are changed
generally tolead these systems into this transitional area between order and chaos. This idea is
very intuitive, since
chaotic dynamical systems are too sensitive to parameter changes, that is, a single mutation leads
the
system into another completely different behavior (sensitive to damage). By contrast, ordered
systems are
more resilient to damage, and a small parameter change will usually result in a small behavior
change
which is ideal for smooth adaptation (hill-climbing) in correlated fitness landscapes. However,
even
though very ordered systems can adapt by accumulation of useful successful variations (because
damage
does not propagate widely), they may not be able 'step out' of certain areas of their fitness
landscapes. It
is here that systems at the edge of chaos enter the scene; they are not as sensitive to damage as
chaotic
systems, but still they are more sensitive than fully ordered systems, and thus, some mutations
will
accumulate (by causing minor changes) and some others will cause major changes in the
dynamics
allowing more distant searches in fitness spaces. These characteristics of simultaneous mutation
buffering
(to small changes) and dramatic alteration of behavior (in response to larger changes) is ideal for
evolvability [Conrad, 1983, 1990].
Chaotic classifications cannot grasp an ordered interaction with an environment, while point
attractors
and simple limit cycles may not allow enough behavior change for a good increase in variety.
The edge of
chaos regime seems to offer a good, intuitive, compromise. However, whatever the regime of a
dynamic
system, self-organization alone cannot escape its own attractor behavior. A given dynamic
system is
always bound to the complexity its attractor landscape allows. For a dynamic system to observe
genuine
emergence of new classifications (conceptual or of functionality) it must change its structure.
Creativity,
or open-ended variety can only be attained by structural perturbation of a dynamical system. One
way or
another, this structural change leading to efficient classification (not just random change), has
only been
achieved through some external influence on the self-organizing system. Artificial neural
networks
discriminate by changing the structure of their connections through an external learning
procedure.
Evolutionary strategies rely on internal random variation (mutation) which must ultimately be
externally
selected. In other words, the self-organizing system must be structurally coupled to some external
system
which acts on structural changes of the first and induces some form of explicit or implicit
selection of its
dynamic representations: selected self-organization.
2.2 Memory and selected self-organization
The dynamical approach of von Foerster [1965] to cognition emphasized the concept of
memory without
a record. By utilizing functionals to change the functions of state-determined systems, von
Foerster
formalized the idea that memory can be observed in systems which are able to change their own
structure
and therefore its dynamics and attractor behavior. Today, we name this kind of memory
distributed, and
the kind of models of memory so attained as connectionist. As previously discussed, for a
self-organizing
system to be informationally open, that is, for it to be able to classify its own interaction with an
environment, it must be able to change its structure, and subsequently its attractor basins,
explicitly or
implicitly. Explicit control of its structure would amount to a choice of a particular dynamics for
a certain
task (the functional would be under direct control of the self-organizing system) and can be
referred to as
learning. Under implicit control, the self-organizing system is subjected to some variation
of its structure
(including its distributed memory) which may or may not be good enough to perform our task.
Those
self-organizing systems which are able to perform the task are thus externally selected by
the environment
to which they are structurally coupled. If reproduction is added to the list of tasks these systems
can
produce based on their dynamic memories, then we have the ingredients for natural selection:
heritable
variation and selection.
This form of situated, embodied, self-organization can be referred to as distributed memory
selected self-
organization. Its relying on some system-environment coupling of structure has been stressed
mostnotably within second-order cybernetics and systems research. Maturana and Varela [1987]
propose
structural coupling as the general mechanism for variety increase, Pask [1976] refers to it as
conversation
in the cognitive realm. Both of these approaches owe a lot to von Foerster's eigenbehavior
notions. More
recently, in the realm of complex systems and evolutionary systems theory, Kauffman [1993] and
others
have relied on the notion of autocatalytic sets which are mutable, heritable, self-replicating, self-
organizing systems evolvable through natural selection.
So far I have maintained that eigenvalues or attractors represent the building blocks of any
system capable
of discriminating its environment through some thus embodied construction. However,
eigenbehavior
(emergent representation) and its variety increase needs a structural coupling of these eigenvalues
with
some externally selective environment. This kind of selected self-organization obliges us "to
understand
perception not just as an interactive dynamical structure, but as a process that arises from a more
fundamental embodiment that makes it possible for evolution to create structures that are
internally
assigned interactive roles. This process carries with it an increase of complexity of the way the
environment is perceived and acted upon" [Etxeberria, 1995]. It also seems to offer a minimum
requirement for evolution and cognitive categorization [Lakoff, 1987; Rocha, 1995d].
Perhaps the most important characteristic of this distributed memory selected self-organization is
the fact
that its specific embodiment both constructs the classification of the environment and ultimately
defines
selection. The consequence of this fact for biological systems is that natural selection (acting on
this form
of self-organization) is not free to evolve any organism, but it is constrained by the
self-organizing
properties of the materiality of the organisms it acts upon _ evolution with both a self-organizing
and
selection component. The consequence for cognitive systems, is that what can be classified is
also
constrained by the particular materiality of the classifying system at stake _ not everything "out
there"
can be grasped. In other words, the particular self-organizing dynamics of a particular classifying
system
constrains the universality of its classification. However, we should look into how can this
process be
made more efficient, and allow for genuine open-ended emergence of variety in
classification.
3 Von Neumann: description based selected evolution
Von Neumann [1966] defended that a threshold of complexity exists, before which complexity
degenerates, and after which complexity can increase in an open-ended fashion. He proposed a
self-replicating scheme based on the notion of a memory-stored description Φ(A) that can be
interpreted by auniversal constructor A to produce A itself. However, to avoid a logical paradox
of self-reference, the
description, which cannot describe itself, must be both copied (uninterpreted role) and
translated
(interpreted role ) into the described automaton. This way, in addition to the universal
constructor, an
automaton B capable of copying any description, Φ, is included in the self-replication
scheme. A third
automaton C is also included to effect all the manipulation of descriptions necessary. To
sum it up, the
self-replicating system contains the set of automata (A + B + C) and a
description Φ(A + B + C); the
description is fed to B which copies it and to A which constructs another
automaton (A + B + C); the copy
is then handled separately to the new automaton which together with this description is also able
to self-
reproduce.
3.1 Descriptions and open-ended evolution
As Von Neumann [1966] discussed, if the description of the self-reproducing automata is
changed
(mutated), in a way as to not affect the basic functioning of (A + B + C)
then, the new automaton (A + B
+ C)' will be slightly different from its parent. Von Neumann used a new automaton
D to be included in
the self-replicating organism, whose function does not disturb the basic performance of
(A + B + C); ifthere is a mutation in the D part of the description,
say D', then the system (A + B + C + D) + Φ(A + B +
C + D') will produce (A + B + C + D') + Φ(A + B + C + D').
Von Neumann [1966, page 86] further
proposed that non-trivial self-reproduction should include this "ability to undergo inheritable
mutations as
well as the ability to make another organism like the original", to distinguish it from "naive" self-
reproduction like growing crystals. Von Neumann's model clearly does not rely on a distributed
but on a
local kind of memory. Descriptions entail a symbol system on which construction commands are
cast.
These commands are not distributed over patterns of activation of the components of a dynamic
system,
but instead localized on "inert" structures which can be used at any time _ a sort of random
access
memory. By "inert" I mean material structures with many dynamically equivalent states, in other
words,
the semantic relation, or what the structures are used to refer to, must possess a large degree of
arbitrariness so that certain representations are not much more probable than others. In the
genetic system,
any sequence of nucleotides is possible, and its informational value is not dependent on the
particular
attractor behavior of DNA or RNA dynamics.
Why then is there an advantage of local memory over distributed memory self-replication? Von
Neumann's argument mainatins that if we do not have symbolic descriptions directing
self-replication,
then an organism must replicate through material self-inspection of its parts. In other words, the
dynamics
must be able to produce copies of itself by template identification of parts existing in its
environment. The
simplest way would be to have every part of the structure individually heritable. Clearly, as
systems grow
in complexity, self-inspection becomes more and more difficult [Pattee, 1995]. The existence of
a
language, a symbol system, allows a much more sophisticated form of communication.
Functional,
dynamic structures do not need to replicate themselves, they are simply constructed from
physically non-
functional (dynamically inert) descriptions. For instance, for an enzyme to replicate itself, it
would need
to have this intrinsic property of self-replication "by default", or it would have to be able to
assemble
itself from a pool of existing parts, but for this, it would have to "unfold" so that its internal parts
could be
reconstituted for the copy to be produced [Pattee, 1995]. With the genetic code, however, none of
these
complicated "gimmicks" are necessary: functional molecules can be simply folded from inert
messages.
This method is by far more general since any functional molecule (with limitations to be
discussed ahead)
can be produced from a description, not merely those that either happen to be able to
self-reproduce, or
those that can unfold and fold at will to be reproduced from available parts. The evolution of
distributed
memory based self-organizing systems is restricted to this type of trivial (in von Neumann's
sense) or
through self-inspection (self-description [Kampis, 1991]) reproduction.
The symbol system, with its utilization of inert structures, opens up a whole new universe of
functionality
which is not available for purely dynamical self-replication. In this sense, it can evolve functions
in an
open-ended fashion. The threshold of complexity proposed by Von Neumann is taken by some
(e.g.
Pattee, Cariani, Kampis in Rocha [1995a]) as another category of self-organization which is
capable of
creative organization and selection from outside. Following our rationale above, we can call it
local
memory selected self-organization, or description based selected self-organization. In
biology, this means
that living systems can follow a largely open-ended evolutionary history (von Neumann;s
threshold of
complexity). In the cognitive realm, the introduction of symbols also opened up a whole new
world of
communication possibilities as the aspects of an environment that can be communicated between
individuals is not restricted to only those things we can "show" or otherwise somehow physically
mimic:
the displacement of local observations.
3.2 Semantic Closure: open-endedness, materiality, and universality
The notion of description implies a self-referential linguistic mechanism. A description must
be cast on
some symbol system while it must also be implemented on some physical structure. Since
manyrealizations of the same symbol system are possible, viewing descriptions only as physical
systems
explains nothing about their symbolic nature in the control of construction. When A
interprets a
description to construct some automaton, a semantic code is utilized to map instructions
into physical
actions to be performed. When B copies a description, only its syntactic aspects
are replicated. Now, the
language of this semantic code presupposes a set of material primitives (e.g. parts and processes)
for
which the instructions are said to "stand for". In other words, descriptions are not universal as
they refer
to some material constituents which cannot be changed without altering the significance of the
descriptions. We can see that a self-reproducing organism following this scheme is an
entanglement of
symbolic controls and material constraints which is closed on its semantics. Howard Pattee
[1982, 1995]
calls such a principle of self-organization semantic closure.
It is important to understand that when we say that a description based selected self-organizing
system is
endowed with open-ended evolutionary potential we do not believe it is universal, that is, that
any
physical system can be evolved. A given semantically closed system is based on some sort of
coding
mechanism between inert and functional structures. However, the code and the associated
construction are
built on some material substrate constraining the whole semantic closure: there is a finite number
of
functional structures which may be constructed with a given set of parts. The degree of
open-endedness
will be dependent on the representational potential of this code. In other words, the larger the
number of
possible equally dynamically inert structures, the larger the universe of functionality that can be
represented in them. For instance, living systems cannot evolve any functional structure
whatsoever (we
have never seen animals on wheels for instance), but still the number of possible functional
combinations
attainable with the DNA-protein code system is very large, far beyond computational limits. In
this sense,
the emergence of functionality is open-ended [Cariani, 1989, 1993] though not universal.
 It is here that the emphasis on the symbolic level of
open-ended evolutionary systems must be tamed. Strong
Darwinism, has emphasized the nature of the symbolic
description of living systems, as much as strong
cognitivism has emphasized the symbolic nature of
cognition. However, semantic closure with its
description based selected self-organization is not
reiterating this position. The symbolic component of
open-ended evolutionary systems is stressed, but the
material, dynamic, self-organizing characteristics of
matter are equally stressed. It is the ultimate inclusive
approach which is neither reductionist nor dualist
[Pattee, 1995]. While it is maintained that a purely
physical description or dynamics will not explain
symbolic function (as several material systems may
implement the same function), it is also maintained that
different material structures will not have identical domains of potentially evolvable functions.
The
important idea is that evolution relies both on self-organization and selection, and only those
self-
organizing systems able to harness their dynamics to obtain a symbolic dimension can have
open-ended
evolutionary potential.
It is here that the emphasis on the symbolic level of
open-ended evolutionary systems must be tamed. Strong
Darwinism, has emphasized the nature of the symbolic
description of living systems, as much as strong
cognitivism has emphasized the symbolic nature of
cognition. However, semantic closure with its
description based selected self-organization is not
reiterating this position. The symbolic component of
open-ended evolutionary systems is stressed, but the
material, dynamic, self-organizing characteristics of
matter are equally stressed. It is the ultimate inclusive
approach which is neither reductionist nor dualist
[Pattee, 1995]. While it is maintained that a purely
physical description or dynamics will not explain
symbolic function (as several material systems may
implement the same function), it is also maintained that
different material structures will not have identical domains of potentially evolvable functions.
The
important idea is that evolution relies both on self-organization and selection, and only those
self-
organizing systems able to harness their dynamics to obtain a symbolic dimension can have
open-ended
evolutionary potential.
4 Evolving semiotics: a conceptual framework for inclusive self-organization
Semiotics concerns the study of signs/symbols in three basic dimensions: syntactics
(rule-basedoperations between signs within the sign system), semantics (relationship between
signs and the world
external to the sign system), and pragmatics (evaluation of the sign system regarding the goals of
their
users) [Morris, 1946]. The importance of this triadic relationship in any sign system has been
repeatedly
stressed by many in the context of biology and genetics [e.g. Waddington, 1972; Pattee, 1982,
1995]; in
particular, Peter Cariani [1995] has presented an excellent discussion of the subject. We can
understand
the semiotics of the genetic system if we consider all processes taking place before translation
(from
transcription to RNA editing) as the set of syntactic operations; the relation between mRNA
(signifier)
and folded amino acid chains (signified), through the genetic code, as the implementation of a
semantic
relation; and finally, the selective pressures on the obtained proteins as the pragmatic evaluation
of the
genetic sign system.
4.1 Semiotics with two symbol types
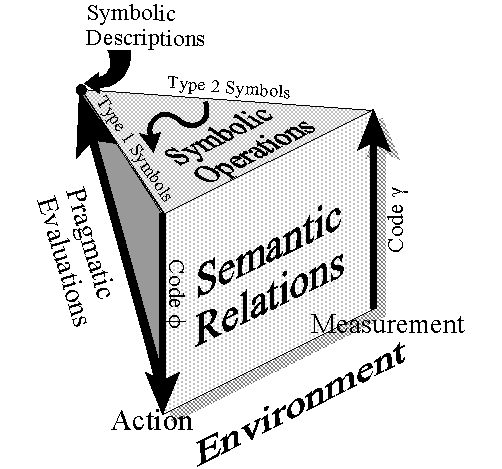
Until now, the semiotics of DNA has been considered strictly
unidirectional: DNA stands for proteins to be constructed. In other
words, the symbolic DNA encodes (through the genetic code) actions
to be performed on some environment. Naturally, through variation
and natural selection (pragmatic evaluations) new semantic relations
are created which are better adapted to a particular environment,
however, real-time contextual measurements are not allowed by this
unidirectional semiotics. If in addition to symbols standing for actions to be
performed, the genetic system is also allowed a second type of symbols
standing for contextual, environmental, measurements, then a richer
semiotics can be created which may have selective advantage in
rapidly changing environments, or in complicated, context
dependent, developmental processes. Figure 2 depicts such a sign
system. The top plane contains two different types of symbols which are combined in different
ways
(symbolic operations). Type 1 symbols stand for actions through a code φ (e.g.
the genetic code) and type
2 symbols stand for measurements through a different code γ which is being
hypothesized here. In Rocha
[1995c] evidence was presented to show that RNA editing may be seen as a mechanism for this
contextual input, at least for certain well known living organisms like the african trypanosomes
[Benne,
1993], and as a potentially important mechanism in the morphogenesis of highly evolved animals
[Lomeli
et al, 1994].
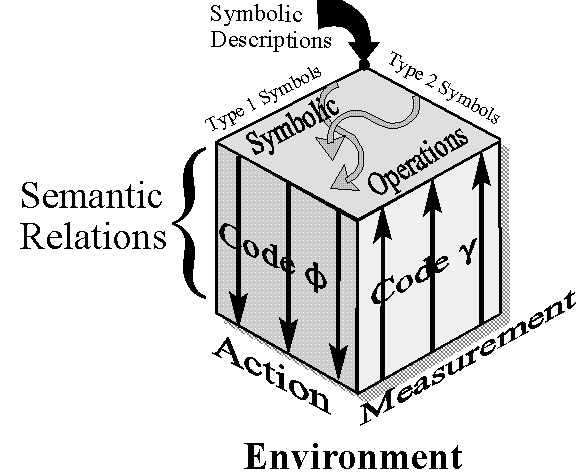 We can think of DNA as a set of symbolic descriptions based on two types of symbols: type
1 symbols are expressed in mRNA molecules and stand for actions to be performed; type
2 symbols are
expressed in some sort editing mechanisms (e.g. gRNA molecules in the genetic system of the
african
trypanosomes) which stand for contextual observables. RNA editing can be
seen as a set of symbolic operations performed with symbols of both
types, resulting in symbols of type 1 to be translated into actions
by the genetic code.
We can think of DNA as a set of symbolic descriptions based on two types of symbols: type
1 symbols are expressed in mRNA molecules and stand for actions to be performed; type
2 symbols are
expressed in some sort editing mechanisms (e.g. gRNA molecules in the genetic system of the
african
trypanosomes) which stand for contextual observables. RNA editing can be
seen as a set of symbolic operations performed with symbols of both
types, resulting in symbols of type 1 to be translated into actions
by the genetic code.
Notice that code γ is proposed here as an abstraction referring to
the set of mechanisms which will link environmental
measurements (context) into type 2 symbols. It is not
expected to function as a proper genetic code. Jon Umerez
[1995] has stressed the importance of a code in any form of
evolving semiotics. In simple terms, what I refer to as a code here
is any mechanism able to relate "inert" material structures to other
material structures with some functional dynamics "by
virtue" of a larger organizational closure. In other words, thefunction of the first material
structures is not dependent on its particular materiality, but on what they are
used to refer to for the imbedding, material, self-referent semantic closure [Pattee, 1995]. Again,
a
semantically closed system, endowed with this kind of symbol/matter code is able establish
open-ended
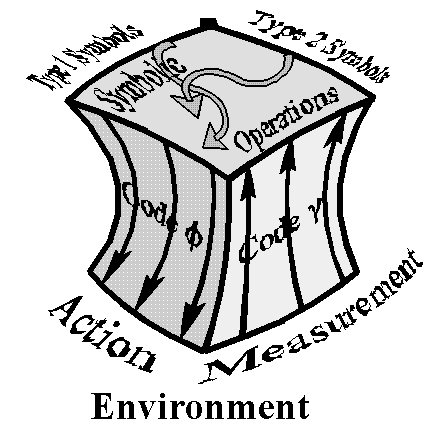
 evolution [Pattee, 1995; Umerez, 1995]. Leaving pragmatic evaluations (selection) out
of the picture momentarily, the semantic closure with two symbol types, which is able
to act as well as perform measurements on its environment can be represented by the
cube in figure 3. The semiotic triadic relationship is only complete when individual
semantic closures are coupled to an environment (measured and acted upon by each
one of them) which ultimately selects (pragmatic evaluation) the most fit amongst
these symbol-matter closures (e.g. in natural selection, those that reproduce the most).
evolution [Pattee, 1995; Umerez, 1995]. Leaving pragmatic evaluations (selection) out
of the picture momentarily, the semantic closure with two symbol types, which is able
to act as well as perform measurements on its environment can be represented by the
cube in figure 3. The semiotic triadic relationship is only complete when individual
semantic closures are coupled to an environment (measured and acted upon by each
one of them) which ultimately selects (pragmatic evaluation) the most fit amongst
these symbol-matter closures (e.g. in natural selection, those that reproduce the most).
4.2 Materiality and implementation dependence: self-organization and selection come
together
The issue of materiality is extremely important for two reasons: (i) all which can be
represented in this
evolutionary semiotics is restricted to what can be constructed by the specific, material,
semantically
closed system in the first place; and (ii) selection is ultimately performed on this specific material
organization capable of performing a number of functions in an environment. The conceptual
framework
put forward by this material, evolutionary, semiotics forces self-organization and selection
together as
two indispensable dimensions of evolutionary systems. Pragmatic
evaluations or selection takes place on particular dynamics, on the other
hand, open-ended evolution is only possible through the existence of a
symbolic dimension mediated through a code. Moreover, this code must be
built out of some materiality which constrains its representation power and
which also ultimately defines eigenbehavior, or an organism's ability to construct
and discriminate its environment. This last point raises the issue of
implementation-independence and multiple realizability [Umerez, 1995].
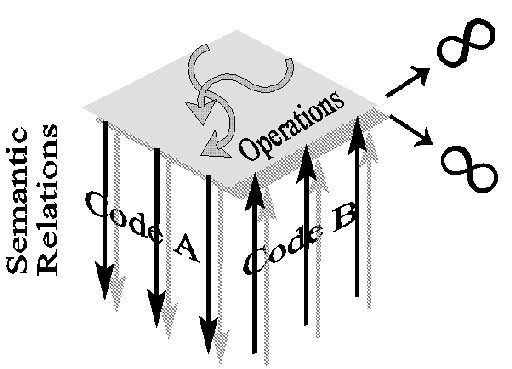 A semantically closed system is not implementation independent because matter
constrains its eigenbehavior as well as its evolutionary potential. The second
constraint is clear when we realize that two distinct closures which at some point establish the
same
eigenbehavior (the same representational function), if materially different, will evolve differently.
The
first constraint is not so clear since we hypothetically allow the idea that two
different closures can have the same representational function. However,
this equivalence can only be established between formal symbol
systems which by definition are not materially constrained and are
therefore universal, that is, the set of possible semantic relations is
infinite (figure 4). Material symbol systems do not have this property. A
coding relation must be formed out of certain available material parts in each
domain (e.g. nucleotides and aminoacids in the genetic code), and no
semantic relation can escape them. In the genetic system we can
represent any protein, but we cannot represent and construct any other
material structure which is not made out of aminoacid chains. Thus, our
semiotics are necessarily constrained by matter, not just due to selection
pressures, but on account of the parts available for the symbol
system itself (figure 5).
A semantically closed system is not implementation independent because matter
constrains its eigenbehavior as well as its evolutionary potential. The second
constraint is clear when we realize that two distinct closures which at some point establish the
same
eigenbehavior (the same representational function), if materially different, will evolve differently.
The
first constraint is not so clear since we hypothetically allow the idea that two
different closures can have the same representational function. However,
this equivalence can only be established between formal symbol
systems which by definition are not materially constrained and are
therefore universal, that is, the set of possible semantic relations is
infinite (figure 4). Material symbol systems do not have this property. A
coding relation must be formed out of certain available material parts in each
domain (e.g. nucleotides and aminoacids in the genetic code), and no
semantic relation can escape them. In the genetic system we can
represent any protein, but we cannot represent and construct any other
material structure which is not made out of aminoacid chains. Thus, our
semiotics are necessarily constrained by matter, not just due to selection
pressures, but on account of the parts available for the symbol
system itself (figure 5).
 Material sign systems are not universal and cannot represent anything whatsoever, but this turns
out to be
their greatest advantage. The price to pay for the universality of formal symbol systems is
complete
specificity, that is, full description of its components and behavior. Conversely, material sign
systems are
built over certain building blocks which do not need a description. For instance, DNA does not
need toencode anything but aminoacid chains, there is no need to include in genetic descriptions
information
regarding the chemical constituents of aminoacids nor instructions on how to fold an aminoacid
chain --
folding comes naturally from the dynamical self-organization of aminoacid chains. Notice how a
logical
simulation of these genetic mechanisms needs to include all this information that comes free
when the
self-organizing characteristics of matter are actually used rather than simulated [Moreno et al,
1994].
Material sign systems are not universal and cannot represent anything whatsoever, but this turns
out to be
their greatest advantage. The price to pay for the universality of formal symbol systems is
complete
specificity, that is, full description of its components and behavior. Conversely, material sign
systems are
built over certain building blocks which do not need a description. For instance, DNA does not
need toencode anything but aminoacid chains, there is no need to include in genetic descriptions
information
regarding the chemical constituents of aminoacids nor instructions on how to fold an aminoacid
chain --
folding comes naturally from the dynamical self-organization of aminoacid chains. Notice how a
logical
simulation of these genetic mechanisms needs to include all this information that comes free
when the
self-organizing characteristics of matter are actually used rather than simulated [Moreno et al,
1994].
5 What does it mean for applications?
5.1 Evolutionary strategies: selection alone
The underlying idea of computational evolutionary strategies (ES) is the separation of
solutions for a
particular problem (e.g. a machine) from descriptions of those solutions through a code.
Genetic
algorithms (GA's) work on these descriptions and not on the solutions themselves, that is,
variation is
applied to descriptions, while the respective solutions are evaluated, and the whole
(description-solution)
selected according to this evaluation. This separation follows von Neumann's self-reproducing
scheme
which is able to increase the complexity of the machines described. This leads to the conclusion
that the
form of organization attained by GA's is not self-organizing in the sense of a boolean network or
cellular
automata. Even though the solutions are obtained from the interaction of a population of
elements, and in
this sense following the general rules usually observed by computationally emergent systems,
they do not
strictly self-organize since they rely on the selective pressures of some fitness
function. The order so
attained is not solely a result of the internal dynamics of a collection of interacting elements, but
also
dictated by the external selection criteria. To say that the populations of descriptions of
solutions self-
organize at all in ES may stretch the concept of self-organization a bit too far. ES rely on
different
concepts: first, with the description-solution dichotomy the concept of local memory is
introduced;
second, the transition rules of ES are not state-determined _ variation is stochastic; third, as
already
discussed, selection is external to the populations of descriptions. This way, we can hardly say
that a
population of memories is interacting with any sort of "self-dynamics": the solutions
reached by a GA do
not self-organize but are a result of external variation and selection. For all these reasons, it is
therefore
natural to think of ES as completely distinct from self-organization. It is perhaps useful to think
of ES as
modeling a very different aspect of biological systems that has to do with natural selection. Self-
organizing systems model the abstract, internal, characteristics of matter, while ES model the
existence
of, external, selective pressures on populations of varying memory based descriptions of some
system.
5.2 Artificial semantic relations: the origin problem
The coded relationship between descriptions and solutions for some task in ES is imposed at
the onset by
the users of such systems. Likewise, the database symbols of some artificial intelligence program
are
externally related to some categories its users are interested in. Both have to do with the issue of
representation in computational domains. All formal systems must have their symbols related to
some
meaning by the external intervention of some user [Rocha, 1995b], in other words, a formal
system
cannot change the signifier/signified primitives imposed when it is started, and create new
observables
[Cariani, 1991]. In the field of GA's some [Mitchell and Forrest, 1994] have been calling for
more
research to be done on schemes that may allow the evolution of the description/solution
relationship
itself, that is, the evolution of a code. The same quest takes place in cognitive science for some
way to
ground the symbols of artificial intelligence models [Harnad, 1990].
Basically, everyone is one way or another dealing with the origin of symbols problem, or in other
words,
the matter/symbol problem. Some explain symbols away by searching explanations in the
dynamics of
cognitive and biological systems [e.g. Churchland and Sejnowski, 1991] while others, usually in
strong
computationalist fields, will look solely at the purely symbolic aspects of complicated systems.
Few havebeen calling for the inclusion of both aspects into complementary approaches [Pattee,
1978; Lakoff, 1987;
Cariani, 1987; Varela, Thompson, and Rosch, 1991; Etxeberria, 1995]. This latter view calls for
an
embodiment of models of life and cognitionin such a way as to be able to study the origin
problems
within an inclusive framework where material and computational aspects are intertwined.
In any case, however far we may be from solving any problems of origin, we may still recognize
that both
life and cognition rely on complementary dynamical and symbolic characteristics. Even if we do
not yet
know how these aspects ever came to be brought together, we should build artificial models
using both of
these aspects (or their simulations) to our advantage, since they have proved to be immensely
powerful
for natural organisms. For instance, in [Rocha, 1995c], even though using a fixed computational
coding
relations between descriptions and solutions in a GA, I proposed the establishment of, stochastic,
contextual constraints on this coding relation following the basic mechanisms of RNA editing
found in a
variety of living organisms. These contextual GA's, though completely computational, are able to
change
the way they produce solutions from the same genetic description, according to changes in their
environments. They are an instance of the two symbol type semiotic model discussed in section
3, and
can be said to evolve an internal control of genetic expression which may be of use for organisms
whose
environment is subjected to cyclic changes.
5.3 Genetic algorithms and development: self-organization and selection in Artificial Life
Lately much attention has been posited on evolutionary strategies that bring together
self-organizing
systems and natural selection inspired algorithms. Particularly in the field of Artificial Life,
Kitano[1994],
and Dellart and Beer [1994], have proposed GA's which do not encode directly their solutions,
but rather
encode generic rules (through L-Systems) which develop into boolean networks simulating given
metabolic cycles. With these approaches, GA's no longer model exclusively selection, but also a
self-
organizing dimension standing for some materiality. The GA does not search the very large space
possible solutions, but a space of basic rules which can be manipulated to build different
self-organizing
networks. These networks are then started (sometimes with some learning algorithm) and will
converge to
some attractor behavior standing for a solution of our simulation. Rather than directly encoding
solutions,
the GA harnesses a space of possible self-organizing networks which will themselves converge
to a
solution -- emergent morphology.
The computational advantage of these systems lies on the tremendous reduction of the
algorithm's search
space since the solutions do not have to be encoded in all details, the emergent morphology
"takes care"
of details we do not need to encode. In particular, I have proposed a developmental scheme
[Rocha,
1995c] which uses the same search space (based on fuzzy rules) for whatever number of
simulation
primitives we desire, in other words, a generic GA which uses the same state space regardless of
the
simulation task by utilizing an emergent morphology scheme based on fuzzy logic. By simulating
both
selection and self-organization, the size of descriptions is dramatically reduced, and an avenue is
opened
for studying the simulation of both the symbolic and material aspects of evolutionary
systems.
5.4 Categorization and constructivism: uncertainty and belief in artificial intelligence
Eleanor Rosch [1978] and George Lakoff [1987], among others, have stressed the
importance of an
embodiment of cognition to deal with its representation issues. In Rocha [1994b, 1995d, 1995e] I
have
introduced a set structure called evidence set based on fuzzy logic and the Dempster-Shafer
[Shafer,
1976] theory of evidence. These structures allow the inclusion of all forms of uncertainty
recognized in
information theory [Klir, 1993] as well as a formalization of belief and contextual dependencies
in a set
format. Evidence sets do not specifically include an account of materiality, however, the
formalization of
belief and context allows larger imbedding models of cognitive categorization to base the belief
andcontextual strengths of concept membership on specific material constraints, or its simulation
through,
say, neural networks.
The contextual pointers of evidence sets are related to Pask's [1976] P-individuals in his
conversation
theory and are thus embedded in a constructivist framework which emphasizes the construction
of a
reality in terms of a cognitive system's specific materiality and environmental coupling. This is
also a
direct result of von Foerster's formulation of eigenbehavior and an aid to establishing another
instance of
the semiotic model of section 3 in the cognitive realm. It can be seen to offer a constructivist
position of
representation which stresses embodiment, but must also, on the other hand, concede that in an
evolutionary context, the construction of categories must have a representational relation to
aspects of the
organism's environment, or its categorization would not have survival value in that particular
organism/environment structural coupling [Medina-Martins and Rocha, 1992; Rocha, 1995d]. In
other
words, embodiment does not eradicate the necessity to still explain some sort of representational
relation
between constructed categories and the cognitive system's context.
6 Conclusions: selection meets self-organization
I have stressed that though self-organizing systems with distributed memory represent a
minimum
requirement for evolutionary systems, their evolutionary potential is much larger, possibly
open-ended, if
further endowed with dynamically "inert" structures to be used by their classification
mechanisms. It was
stressed that this by no means entails a return to purely symbolic approaches to cognition, nor a
belief in
the absolute sovereignty of natural selection in evolution. Rather, it is a call for more inclusive,
hybrid
approaches to such evolutionary and learning systems. In artificial Life this implies building
models
which bring together self-organizing mechanisms, such as cellular automata or boolean networks,
with
genetic algorithms (with varying degrees of control of their genetic expression). In Artificial
Intelligence
it implies the establishment of models able to go beyond connectionist classification, by
inclusion of
higher level accounts of cognitive categorization.
7 References
Benne, Rob (Ed.) [1993]. RNA Editing: The Alteration of Protein Coding Sequences of
RNA. Ellis Horwood.
Cariani, Peter [1991]."Some
Epistemological Implications of Devices which Construct their own Sensors and
Effectors." In: Towards a Practice of Autonomous Systems. Varela and Bourgine
(Eds.). MIT pp 484-493.
Cariani, Peter [1995]."Towards an evolutionary semiotics: the role of
symbols in organisms and adaptive devices."
In: Proc. of the ISES Vienna 1995. Stanley Salthe and Gertrudis Van de Vijver (eds).
In Press.
Churchland, Patricia S. and T.J.
Sejnowski [1992]. The Computational Brain. MIT Press.
Conrad, Michael [1983].
Adaptability. Plenum Press.
Conrad, Michael [1990]."The
geometry of evolutions." In: BioSystems Vol. 24, pp. 61-81.
Dellaert, F. and R.D. Beer
[1994]."Toward an evolvable model of development for autonomous agent
synthesis." In:
Artificial Life IV. R. Brooks and P. Maes (Eds.). MIT Press.
Etxeberria, Arantza
[1995]."Embodiment of Natural and Artificial Systems." In: Proceedings of the
International
Seminar on Evolutionary Systems, Vienna 1995. G. van de Vijver and S. Salthe (Eds.).
In Press.
Haken, H. [1977]. Synergetics.
Springer-Verlag.
Harnad, Stevan [1990]."The
symbol grounding problem." In: Emergent Computation. S. Forrest (Ed.). MIT
Press.
Kampis, George [1991].
Self-Modifying Systems in Biology and Cognitive Science. .
Kauffman, S. [1993]. The Origins
of Order: Self-Organization and Selection in Evolution. Oxford University Press.
Kitano, Hiroaki
[1994]."Evolution of Metabolism for Morphogenesis." In: Artificial Life IV.
Brooks, R.A. and P.
Maes. MIT Press. pp 49-58..
Klir, George J. [1991]. Facets of
Systems Science. Plenum Press.
Klir, George J.
[1993]."Developments in uncertainty-based information." In: Advances in
Computers. M. Yovits(Ed.). Vol. 36, pp 255-332.
Lakoff, G. [1987] Women, Fire, and Dangerous Things. Chicago University Press.
Maturana, H., and F. Varela [1987].
The Tree of Knowledge. New Science Library.
Medina-Martins, Pedro R. and Luis
Rocha [1992]."The in and the out: an evolutionary approach." In: Cybernetics
and Systems Research '92. Robert Trappl. World Scientific Press. pp 681-689.
Mitchell, M. and S. Forrest
[1994]."Genetic algorithms and Artificial Life." In: Artificial Life Vol. 1, pp
267-289.
Moreno, A., A. Etxeberria, and J.
Umerez [1994]."Universality without matter?." In: Artificial Life IV. R.
Brooks
and P. Maes (Eds). MIT Press, pp.406-410.
Morris, Charles W. [1946]. Signs,
Language, and Behavior. G. Braziller, New York.
Nicolis, G. and I. Prigogine [1977].
Self-Organization in Nonequilibrium Systems
Packard, N. [1988] "Adaptation to the
edge of chaos". In: Complexity in Biologic Modelling. Kelso and Shlesinger
Pask, Gordon [1976].
Conversation Theory: Applications in Education and Epistemology. Elsevier.
Pask, Gordon [1992]."Different
kinds of Cybernetics." In: New Perspectives on Cybernetics. Gertrudis Van de
Vijver (Ed.). Kluwer Academic Publishers. pp. 11-31.
Pattee, Howard H. [1978]."The
complementarity principle in biological and social structures." In: Journal of Social
and Biological Structures Vol. 1, pp. 191-200.
Pattee, Howard H. [1982]."Cell
psychology: an evolutionary approach to the symbol-matter problem." In: Cognition
and Brain Theory Vol. 5, no. 4, pp 325-341.
Pattee, Howard H.
[1995]."Evolving self-seference: matter, symbols, and semantic closure."
Communication and
Cognition - AI Vol. 12, nos 1-2 (Rocha [1995a]). pp. 9-27 .
Prigogine, I. [1985]."New perspectives on complexity." In:
The Science and Praxis of Complexity. United Nations
Library. Reprinted in Klir [1991], pp. 483-492.
Rocha, Luis M. [1994a]."Von
Foerster's cognitive tiles: semantically closed building blocks for AI and Alife." In:
Cybernetics and Systems ' 94. Robert Trappl (Ed.). Vol. 1, pp 621-628. World
Scientific Press.
Rocha, Luis M.
[1994b]."Cognitive Categorization revisited: extending interval valued fuzzy sets as
simulation tools
for concept combination." In: Proc. of the 1994 Int. Conf. of NAFIPS/IFIS/NASA.
IEEE. pp 400-404.
Rocha, Luis M. (Ed.) [1995a].
special issue in Self-Reference in Biological and Cognitive Systems. Communication
and Cognition - Artificial Intelligence Vol. 12, nos. 1-2
Rocha, Luis M.
[1995b]."Artificial semantically closed objects." Communication and Cognition -
AI Vol. 12, nos. 1-
2 (Rocha[1995a]). pp. 63-90.
Rocha, Luis M. [1995c]."Contextual Genetic Algorithms: Evolving Developmental
Rules." In: Advances in
Artificial Life. F. Moran, A. Moreno, J.J. Merelo, and P. Chacon (Eds.). Springer-Verlag.
pp. 368-382.
Rocha, Luis M.
[1995d]."(Interval based) Evidence Sets: Modelling Subjective Categories." In:
International
Journal of General Systems In Press.
Rocha, Luis M.
[1995e]."Computing Uncertainty in Interval Based Sets." In: Applications of
Interval Computation.
R.B. Kearfott and V. Kreinovich (Eds.). Kluwer. (In Press).
Rosch, E. [1978]."Principles of
Categorization." Cognition and Categorization. Rosch and Lloyd
(Eds.).Hillsdale.




